Cerebellum
|
WikiDoc Resources for Cerebellum |
|
Articles |
|---|
|
Most recent articles on Cerebellum |
|
Media |
|
Evidence Based Medicine |
|
Clinical Trials |
|
Ongoing Trials on Cerebellum at Clinical Trials.gov Clinical Trials on Cerebellum at Google
|
|
Guidelines / Policies / Govt |
|
US National Guidelines Clearinghouse on Cerebellum
|
|
Books |
|
News |
|
Commentary |
|
Definitions |
|
Patient Resources / Community |
|
Patient resources on Cerebellum Discussion groups on Cerebellum Patient Handouts on Cerebellum Directions to Hospitals Treating Cerebellum Risk calculators and risk factors for Cerebellum
|
|
Healthcare Provider Resources |
|
Causes & Risk Factors for Cerebellum |
|
Continuing Medical Education (CME) |
|
International |
|
|
|
Business |
|
Experimental / Informatics |
Editor-In-Chief: C. Michael Gibson, M.S., M.D. [1]
Overview
The cerebellum (Latin: "little brain") is a region of the brain that plays an important role in the integration of sensory perception and motor output. Many neural pathways link the cerebellum with the motor cortex—which sends information to the muscles causing them to move—and the spinocerebellar tract—which provides feedback on the position of the body in space (proprioception). The cerebellum integrates these pathways, using the constant feedback on body position to fine-tune motor movements.
Because of this 'updating' function of the cerebellum, lesions within it are not so debilitating as to cause paralysis, but rather present as feedback deficits resulting in disorders in fine movement, equilibrium, posture, and motor learning. Initial observations by physiologists during the 18th century indicated that patients with cerebellar damage show problems with motor coordination and movement. Research into cerebellar function during the early to mid 19th century was done via lesion and ablation studies in animals. Research physiologists noted that such lesions led to animals with strange movements, awkward gait, and muscular weakness. These observations and studies led to the conclusion that the cerebellum was a motor control structure.[1] However, modern research shows that the cerebellum has a broader role in a number of key cognitive functions, including attention and the processing of language, music, and other sensory temporal stimuli.
General features
The cerebellum is located in the inferior posterior portion of the head (the hindbrain), directly dorsal to the pons, and inferior to the occipital lobe (Figs. 1 and 3). Because of its large number of tiny granule cells, the cerebellum contains nearly 50% of all neurons in the brain, but it only takes up 10% of total brain volume. The cerebellum receives nearly 200 million input fibers; in contrast, the optic nerve is composed of a mere one million fibers.
The cerebellum is divided into two large hemispheres, much like the cerebrum, and contains ten smaller lobules. The cytoarchitecture (cellular organization) of the cerebellum is highly uniform, with connections organized into a rough, three-dimensional array of perpendicular circuit elements. This organizational uniformity makes the nerve circuitry relatively easy to study. To envision this "perpendicular array," one might imagine a tree-lined street with wires running straight through the branches of one tree to the next.
Development and evolution

During the early stages of embryonic development, the brain starts to form in three distinct segments: the prosencephalon, mesencephalon, and rhombencephalon. The rhombencephalon is the most caudal (toward the tail) segment of the embryonic brain; it is from this segment that the cerebellum develops. Along the embryonic rhombencephalic segment develop eight swellings, called rhombomeres. The cerebellum arises from two rhombomeres located in the alar plate of the neural tube, a structure that eventually forms the brain and spinal cord. The specific rhombomeres from which the cerebellum forms are rhombomere 1 (Rh.1) caudally (near the tail) and the "isthmus" rostrally (near the front).[2]
Two primary regions are thought to give rise to the neurons that make up the cerebellum. The first region is the ventricular zone in the roof of the fourth ventricle. This area produces Purkinje cells and deep cerebellar nuclear neurons. These cells are the primary output neurons of the cerebellar cortex and cerebellum. The second germinal zone (cellular birthplace) is known as the external granular layer. This layer of cells—found on the exterior the cerebellum—produces the granule neurons. Once born, the granule neurons migrate from this exterior layer to form an inner layer known as the internal granule layer. The external granular layer ceases to exist in the mature cerebellum, leaving only granule cells in the internal granule layer. The cerebellar white matter may be a third germinal zone in the cerebellum; however, its function as a germinal zone is controversial.
The cerebellum is of archipalliar phylogenetic origin. The pallium is a term for gray matter that forms the cortex. The archipallium is the one of the most evolutionarily primitive brain regions. The circuits in the cerebellar cortex look similar across all classes of vertebrates, including fish, reptiles, birds, and mammals (e.g., Fig. 2). This has been taken as evidence that the cerebellum performs functions important to all vertebrate species.
Anatomy
The cerebellum contains similar gray and white matter divisions as the cerebrum. Embedded within the white matter—which is known as the arbor vitae (Tree of Life) in the cerebellum due to its branched, treelike appearance—are four deep cerebellar nuclei. Three gross phylogenetic segments are largely grouped by general function. The three cortical layers contain various cellular types that often create various feedback and feedforward loops. Oxygenated blood is supplied by three arterial branches off the basilar and vertebral arteries.
Divisions
The cerebellum can be divided according to three different criteria: gross anatomical, phyologenetical, and functional.
Gross anatomical divisions
On gross inspection, three lobes can be distinguished in the cerebellum: the flocculonodular lobe, the anterior lobe (rostral to the "primary fissure"), and the posterior lobe (dorsal to the "primary fissure"). The latter two can be further divided in a midline cerebellar vermis and lateral cerebellar hemispheres.
 |
 |
Phylogenetic and functional divisions
The cerebellum can also be divided in three parts based on both phylogenetic criteria (the evolutionary age of each part) and on functional criteria (the incoming and outgoing connections each part has and the role played in normal cerebellar function). From the phylogenetically oldest to the newest, the three parts are:
| Functional denomination (phylogenetic denomination) | Anatomical parts | Role |
| Vestibulocerebellum (Archicerebellum) | Flocculonodular lobe (and immediately adjacent vermis) | The vestibulocerebellum regulates balance and eye movements. It receives vestibular input from both the semicircular canals and from the vestibular nuclei, and sends fibres back to the medial and lateral vestibular nuclei. It also receives visual input from the superior colliculi and from the visual cortex (the latter via the pontine nuclei, forming a cortico-ponto-cerebellar pathway). Lesions of the vestibulocerebellum cause disturbances of balance and gait. |
| Spinocerebellum (Paleocerebellum) | Vermis and intermediate parts of the hemispheres ("paravermis") | The spinocerebellum regulates body and limb movements. It receives proprioception input from the dorsal columns of the spinal cord (including the spinocerebellar tract) as well as from the trigeminal nerve, as well as from visual and auditory systems. It sends fibres to deep cerebellar nuclei which in turn project to both the cerebral cortex and the brain stem, thus providing modulation of descending motor systems. The spinocerebellum contains sensory maps as it receives data on the position of various body parts in space: in particular, the vermis receives fibres from the trunk and proximal portions of limbs, while the intermediate parts of the hemispheres receive fibres from the distal portions of limbs. The spinocerebellum is able to elaborate proprioceptive input in order to anticipate the future position of a body part during the course of a movement, in a "feed forward" manner. |
| Cerebrocerebellum (Neocerebellum) | Lateral parts of the hemispheres | The neocerebellum is involved in planning movement and evaluating sensory information for action. It receives input exclusively from the cerebral cortex (especially the parietal lobe) via the pontine nuclei (forming cortico-ponto-cerebellar pathways), and sends fibres mainly to the ventrolateral thalamus (in turn connected to motor areas of the premotor cortex and primary motor area of the cerebral cortex) and to the red nucleus (in turn connected to the inferior olivary nucleus, which links back to the cerebellar hemispheres). The neocerebellum is involved in planning movement that is about to occur[3] and has purely cognitive functions as well. |
Much of what is understood about the functions of the cerebellum stems from careful documentation of the effects of focal lesions in human patients who have suffered from injury or disease or through animal lesion research.
Deep nuclei
The four deep cerebellar nuclei are in the center of the cerebellum, embedded in the white matter. These nuclei receive inhibitory (GABAergic) inputs from Purkinje cells in the cerebellar cortex and excitatory (glutamatergic) inputs from mossy fiber pathways. Most output fibers of the cerebellum originate from these nuclei. One exception is that fibers from the flocculonodular lobe synapse directly on vestibular nuclei without first passing through the deep cerebellar nuclei. The vestibular nuclei in the brainstem are analogous structures to the deep nuclei, since they receive both mossy fiber and Purkinje cell inputs.
From lateral to medial, the four deep cerebellar nuclei are the dentate, emboliform, globose, and fastigial. An easy mnemonic device to remember these names and positions relative to their position from the midline is the phrase "Don't Eat Greasy Food", where each letter indicates the lateral to medial location in the cerebellar white matter. Some animals do not have distinct emboliform and globose nuclei, instead having a single, fused nucleus interpositus (interposed nucleus). In animals with distinct emboliform and globose nuclei, the term interposed nucleus is often used to refer collectively to these two nuclei.
In general, each pair of deep nuclei is associated with a corresponding region of cerebellar surface anatomy. The dentate nuclei are deep within the lateral hemispheres, the interposed nuclei are located in the paravermal (intermediate) zone, and the fastigial nuclei are in the vermis. These structural relationships are generally maintained in the neuronal connections between the nuclei and associated cerebellar cortex, with the dentate nucleus receiving most of its connections from the lateral hemispheres, the interposed nuclei receiving inputs mostly from the paravermis, and the fastigial nucleus receiving primarily afferents from the vermis.
Cortical layers


There are three layers to the cerebellar cortex; from outer to inner layer, these are the molecular, Purkinje, and granular layers. The function of the cerebellar cortex is essentially to modulate information flowing through the deep nuclei. The microcircuitry of the cerebellum is schematized in Figure 5. Mossy and climbing fibers carry sensorimotor information into the deep nuclei, which in turn pass it on to various premotor areas, thus regulating the gain and timing of motor actions. Mossy and climbing fibers also feed this information into the cerebellar cortex, which performs various computations, resulting in the regulation of Purkinje cell firing. Purkinje neurons feed back into the deep nuclei via a potent inhibitory synapse. This synapse regulates the extent to which mossy and climbing fibers activate the deep nuclei, and thus control the ultimate effect of the cerebellum on motor function. The synaptic strength of almost every synapse in the cerebellar cortex has been shown to undergo synaptic plasticity. This allows the circuitry of the cerebellar cortex to continuously adjust and fine-tune the output of the cerebellum, forming the basis of some types of motor learning and coordination. Each layer in the cerebellar cortex contains the various cell types that comprise this circuitry.
Granular layer
The innermost layer contains the cell bodies of two types of cells: the numerous and tiny granule cells, and the larger Golgi cells. Mossy fibers enter the granular layer from their main point of origin, the pontine nuclei. These fibers form excitatory synapses with the granule cells and the cells of the deep cerebellar nuclei. The granule cells send their T-shaped axons—known as parallel fibers—up into the superficial molecular layer, where they form hundreds of thousands of synapses with Purkinje cell dendrites. The human cerebellum contains on the order of 60 to 80 billion granule cells, making this single cell type by far the most numerous neuron in the brain (roughly 70% of all neurons in the brain and spinal cord, combined). Golgi cells provide inhibitory feedback to granule cells, forming a synapse with them and projecting an axon into the molecular layer.
Purkinje layer
The middle layer contains only one type of cell body—that of the large Purkinje cell. Purkinje cells are the primary integrative neurons of the cerebellar cortex and provide its sole output. Purkinje cell dendrites are large arbors with hundreds of spiny branches reaching up into the molecular layer (Fig. 6). These dendritic arbors are flat—nearly all of them lie in planes—with neighboring Purkinje arbors in parallel planes. Each parallel fiber from the granule cells runs orthogonally through these arbors, like a wire passing through many layers. Purkinje neurons are GABAergic—meaning they have inhibitory synapses—with the neurons of the deep cerebellar and vestibular nuclei in the brainstem. Each Purkinje cell receives excitatory input from 100,000 to 200,000 parallel fibers. Parallel fibers are said to be responsible for the simple (all or nothing, amplitude invariant) spiking of the Purkinje cell.
Purkinje cells also receive input from the inferior olivary nucleus via climbing fibers. A good mnemonic for this interaction is the phrase "climb the other olive tree", given that climbing fibers originate from the contralateral inferior olive. In striking contrast to the 100,000-plus inputs from parallel fibers, each Purkinje cell receives input from exactly one climbing fiber; but this single fiber "climbs" the dendrites of the Purkinje cell, winding around them and making a large number of synapses as it goes. The net input is so strong that a single action potential from a climbing fiber is capable of producing a "complex spike" in the Purkinje cell: a burst of several spikes in a row, with diminishing amplitude, followed by a pause during which simple spikes are suppressed.
Molecular layer
This outermost layer of the cerebellar cortex contains two types of inhibitory interneurons: the stellate and basket cells. It also contains the dendritic arbors of Purkinje neurons and parallel fiber tracts from the granule cells. Both stellate and basket cells form GABAergic synapses onto Purkinje cell dendrites.
Peduncles
Similarly, the cerebellum follows the trend of "threes", with three major input and output peduncles (fiber bundles). These are the superior (brachium conjunctivum), middle (brachium pontis), and inferior (restiform body) cerebellar peduncles.
| Peduncle | Description |
| Superior | While there are some afferent fibers from the anterior spinocerebellar tract that are conveyed to the anterior cerebellar lobe via this peduncle, most of the fibers are efferents. Thus, the superior cerebellar peduncle is the major output pathway of the cerebellum. Most of the efferent fibers originate within the dentate nucleus which in turn project to various midbrain structures including the red nucleus, the ventral lateral/ventral anterior nucleus of the thalamus, and the medulla. The dentatorubrothalamocortical (dentate nucleus > red nucleus > thalamus > premotor cortex) and cerebellothalamocortical (cerebellum > thalamus > premotor cortex) pathways are two major pathways that pass through this peduncle and are important in motor planning. |
| Middle | This is composed entirely of afferent fibers originating within the pontine nuclei as part of the massive corticopontocerebellar tract (cerebral cortex > pons > cerebellum). These fibers descend from the sensory and motor areas of the cerebral neocortex and make the middle cerebellar peduncle the largest of the three cerebellar peduncles. |
| Inferior | This carries many types of input and output fibers that are mainly concerned with integrating proprioceptive sensory input with motor vestibular functions such as balance and posture maintenance. Proprioceptive information from the body is carried to the cerebellum via the dorsal spinocerebellar tract. This tract passes through the inferior cerebellar peduncle and synapses within the paleocerebellum. Vestibular information projects onto the archicerebellum. The climbing fibers of the inferior olive run through the inferior cerebellar peduncle. This peduncle also carries information directly from the Purkinje cells out to the vestibular nuclei in the dorsal brainstem located at the junction between the pons and medulla. |
There are three sources of input to the cerebellum, in two categories consisting of mossy and climbing fibers, respectively. Mossy fibers can originate from the pontine nuclei, which are clusters of neurons located in the pons that carry information from the contralateral cerebral cortex. They may also arise within the spinocerebellar tract whose origin is located in the ipsilateral spinal cord. Most of the output from the cerebellum initially synapses onto the deep cerebellar nuclei before exiting via the three peduncles. The most notable exception is the direct inhibition of the vestibular nuclei by Purkinje cells.
Blood supply

Three arteries supply blood to the cerebellum (Fig. 7): the superior cerebellar artery (SCA), anterior inferior cerebellar artery (AICA), and posterior inferior cerebellar artery (PICA).
Superior cerebellar artery
The SCA branches off the lateral portion of the basilar artery, just inferior to its bifurcation into the posterior cerebral artery. Here it wraps posteriorly around the pons (to which it also supplies blood) before reaching the cerebellum. The SCA supplies blood to most of the cerebellar cortex, the cerebellar nuclei, and the middle and superior cerebellar peduncles.
Anterior inferior cerebellar artery
The AICA branches off the lateral portion of the basilar artery, just superior to the junction of the vertebral arteries. From its origin, it branches along the inferior portion of the pons at the cerebellopontine angle before reaching the cerebellum. This artery supplies blood to the anterior portion of the inferior cerebellum, and to the facial (CN VII) and vestibulocochlear nerves (CN VIII).
Obstruction of the AICA can cause paresis, paralysis, and loss of sensation in the face; it can also cause hearing impairment. Moreover, it could cause an infarct of the cerebellopontine angle. This could lead to hyperacusia (dysfunction of the stapedius muscle, innervated by CN VII) and vertigo (wrong interpretation from the vestibular semi-circular canal's endolymph acceleration caused by alteration of CN VIII).
Posterior inferior cerebellar artery
The PICA branches off the lateral portion of the vertebral arteries just inferior to their junction with the basilar artery. Before reaching the inferior surface of the cerebellum, the PICA sends branches into the medulla, supplying blood to several cranial nerve nuclei. In the cerebellum, the PICA supplies blood to the posterior inferior portion of the cerebellum, the inferior cerebellar peduncle, the nucleus ambiguus, the vagus motor nucleus, the spinal trigeminal nucleus, the solitary nucleus, and the vestibulocochlear nuclei.
Dysfunction
Ataxia is a complex of symptoms, generally involving a lack of coordination, that is often found in disease processes affecting the cerebellum. To identify cerebellar problems, the neurological examination includes assessment of gait (a broad-based gait being indicative of ataxia), finger-pointing tests and assessment of posture.[1] Structural abnormalities of the cerebellum (hemorrhage, infarction, neoplasm, degeneration) may be identified on cross-sectional imaging. Magnetic resonance imaging is the modality of choice, as computed tomography is insufficiently sensitive for detecting structural abnormalities of the cerebellum.[4]
Theories about cerebellar function
Two main theories address the function of the cerebellum, both dealing with motor coordination. One claims that the cerebellum functions as a regulator of the "timing of movements". This has emerged from studies of patients whose timed movements are disrupted.[5]
The second, "Tensor Network Theory" provides a mathematical model of transformation of sensory (covariant) space-time coordinates into motor (contravariant) coordinates by cerebellar neuronal networks.[6][7]
Like many controversies in the physical sciences, there is evidence supporting each of the above hypotheses. Studies of motor learning in the vestibulo-ocular reflex and eyeblink conditioning demonstrate that the timing and amplitude of learned movements are encoded by the cerebellum.[8] Many synaptic plasticity mechanisms have been found throughout the cerebellum. The Marr-Albus model mostly attributes motor learning to a single plasticity mechanism: the long-term depression of parallel fiber synapses. The Tensor Network Theory of sensorimotor transformations by the cerebellum has also been experimentally supported.[9][10]
With the advent of more sophisticated neuroimaging techniques such as positron emission tomography (PET),[11] and fMRI,[12] numerous diverse functions are now at least partially attributed to the cerebellum. What was once thought to be primarily a motor/sensory integration region is now proving to be involved in many diverse cognitive functions. Paradoxically, despite the importance of this region and the heterogeneous role it plays in motor and sensory functions, people who have lost their entire cerebellum through disease, injury, or surgery can live reasonably normal lives.
Cerebellar modeling
As mentioned in the preceding section, there have been many attempts to model the cerebellar function.[13] The insights provided by the models have also led to extrapolations in the domains of artificial intelligence methodologies, especially neural networks. Some of the notable achievements have been Cerebellatron ,[14] Cerebellar Model Associative Memory or CMAC networks, and SpikeFORCE for robotic movement control,[15] and "Tensor Network Theory".[16]
Additional images
-
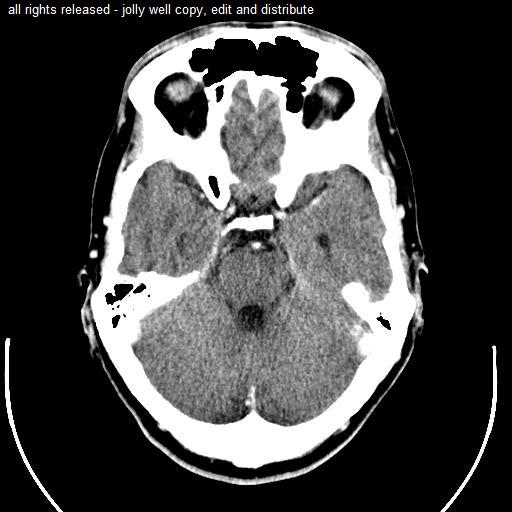
Computed tomography of head, with cerebellum visible at lower part.
-
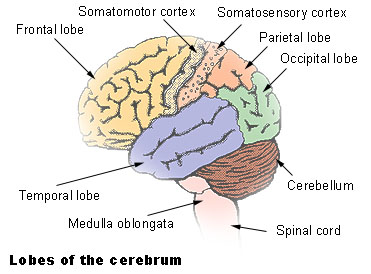
Lobes
-
-
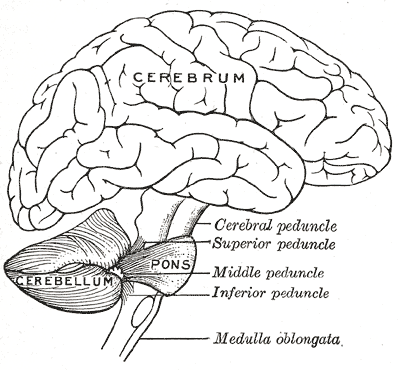
Scheme showing the connections of the several parts of the brain.
-
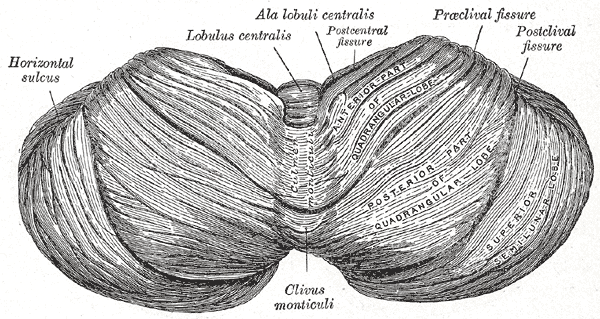
Upper surface of the cerebellum.
-
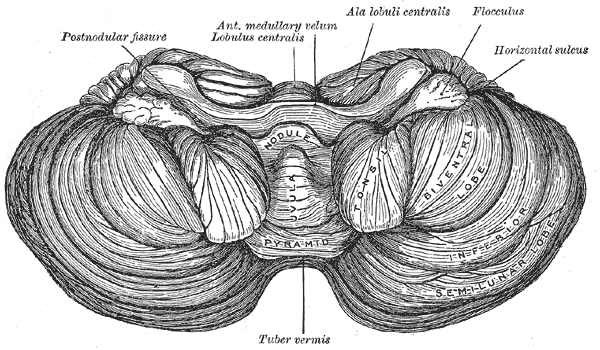
Under surface of the cerebellum.
-
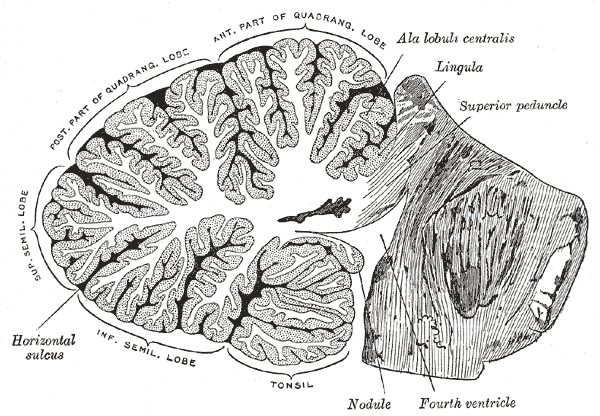
Sagittal section of the cerebellum, near the junction of the vermis with the hemisphere.
-
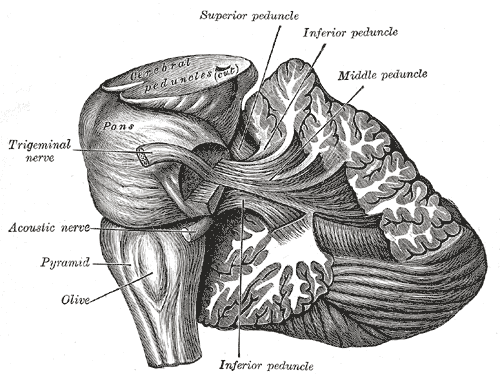
Dissection showing the projection fibers of the cerebellum.
-
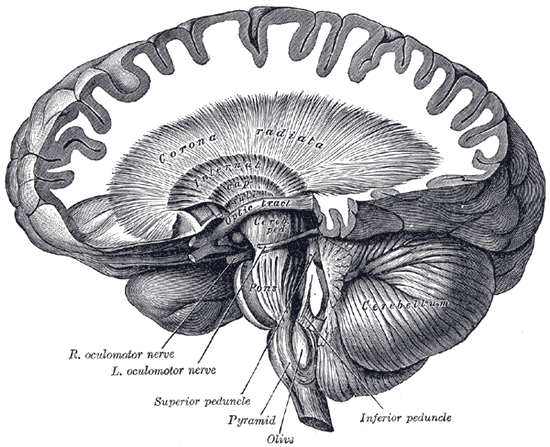
Dissection showing the course of the cerebrospinal fibers.
-
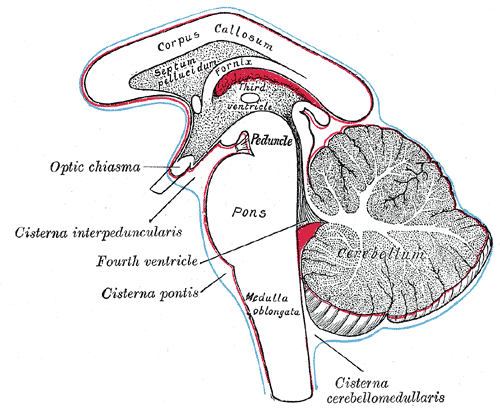
Diagram showing the positions of the three principal subarachnoid cisternæ.
-
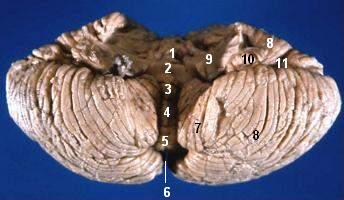
Human cerebellum anterior view
-
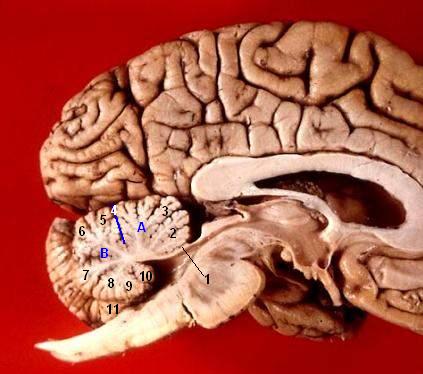
Human brain midsagittal view












See also
External links
- A worldwide list of laboratories that do Template:Wayback
- Cerebellum and timing at University of California, Berkeley
- Cerebellum and multiple sclerosis at mult-sclerosis.org
- Basal ganglia and cerebellum at Washington University in St. Louis
- Cerebellum images at About.com
- Template:BrainMaps
- Histological section of primate cerebellum at University of Birmingham
Further reading
- Ito M. Cerebellum and Neural Control. New York: Raven Press; 1984. ISBN 0-89004-106-7
- Kandel ER, Schwartz JH, Jessell TM. Principles of Neural Science, 4th ed. McGraw-Hill, New York (2000). ISBN 0-8385-7701-6
- Llinás, R, Sotelo C. The Cerebellum Revisited. New York: Springer; 1992. ISBN 0-387-97693-0
- Parent A, Carpenter MB. Carpenter's Human Neuroanatomy. 9th ed. Philadelphia: Williams and Wilkins; 1995. ISBN 0-683-06752-4
References
- ↑ 1.0 1.1 Fine EJ, Ionita CC, Lohr L (2002). "The history of the development of the cerebellar examination". Semin Neurol. 22 (4): 375–84. PMID 12539058.
- ↑ Muller F, O'Rahilly R (1990). "The human brain at stages 21–23, with particular reference to the cerebral cortical plate and to the development of the cerebellum". Anat Embryol (Berl). 182 (4): 375–400. PMID 2252222.
- ↑ Kingsley, R. E. (2000). Consise Text of Neuroscience (2nd edition ed.). Lippincott Williams and Wilkins. ISBN 0-683-30460-7.
- ↑ Gilman S (1998). "Imaging the brain. Second of two parts". N. Engl. J. Med. 338 (13): 889–96. PMID 9516225.
- ↑ Ivry RB, Keele SW, Diener HC (1988). "Dissociation of the lateral and medial cerebellum in movement timing and movement execution". Exp Brain Res. 73 (1): 167–80. PMID 3208855.
- ↑ Pellionisz, A., Llinás, R. (1980). "Tensorial Approach To The Geometry Of Brain Function: Cerebellar Coordination Via A Metric Tensor". Neuroscience. 5: 1125&mdash, -1136.
- ↑ Pellionisz, A., Llinás, R. (1985). "Tensor Network Theory Of The Metaorganization Of Functional Geometries In The Central Nervous System". Neuroscience. 16 (2): 245–273.
- ↑ Boyden ES, Katoh A, Raymond JL (2004). "Cerebellum-dependent learning: the role of multiple plasticity mechanisms". Annu Rev Neurosci. 27: 581–609. PMID 15217344.
- ↑ Pellionisz A, Llinas R (1982). "Space-time representation in the brain. the cerebellum as a predictive space-time metric tensor". Neuroscience. 7 (12): 2949–70. PMID 7162624.
- ↑ Gielen CC, van Zuylen EJ (1986). "Coordination of arm muscles during flexion and supination: application of the tensor analysis approach". Neuroscience. 17 (3): 527–39. PMID 3703248.
- ↑ Obayashi S (2004). "Possible mechanism for transfer of motor skill learning: implication of the cerebellum". Cerebellum. 3 (4): 204–11. PMID 15686098.
- ↑ Kim SG, Ugurbil K, Strick PL (1994). "Activation of a cerebellar output nucleus during cognitive processing". Science. 265 (5174): 949–51. PMID 8052851.
- ↑ Pellionisz, A (1986). "David Marr's Theory of the Cerebellar Cortex: A Model in Brain Theory for the 'Galilean Combination of Simplification, Unification and Mathematization'". In Palm G., & Aertsen, A. Brain Theory. Springer Verlag. pp. 253–257.
- ↑ Dr Evor L. Hines. "Intelligent Systems Engineering - Major Characteristics of Expert Systems and Artificial Neural Networks". Intelligent Systems Engineering (ES3770) Lecture Notes. Retrieved 2006-03-28.
- ↑ "SpikeFORCE: Real-time Spiking Networks for Robot Control" (PDF). Retrieved 2006-03-28.
- ↑ Pellionisz, András J. "Tensor Network Theory of the Central Nervous System". In George Adelman. Encyclopedia of Neuroscience (II ed.). Birkhauser. pp. 1196–1198.
Template:Rhombencephalon
cs:Mozeček
da:Lillehjerne
de:Kleinhirn
eo:Cerebelo
io:Cerebelo
it:Cervelletto
he:צרבלום
la:Cerebellum
lt:Smegenėlės
hu:Kisagy
nl:Kleine hersenen
no:Lillehjernen
simple:Cerebellum
sk:Mozoček
sr:Мали мозак
fi:Pikkuaivot
sv:Lillhjärnan